Medicinsk expert av artikeln

Nya publikationer
äggstock
Senast recenserade: 23.04.2024

Allt iLive-innehåll är mediekontrollerat eller faktiskt kontrollerat för att säkerställa så mycket faktuell noggrannhet som möjligt.
Vi har strikta sourcing riktlinjer och endast länk till välrenommerade media webbplatser, akademiska forskningsinstitut och, när det är möjligt, medicinsk peer granskad studier. Observera att siffrorna inom parentes ([1], [2] etc.) är klickbara länkar till dessa studier.
Om du anser att något av vårt innehåll är felaktigt, omodernt eller på annat sätt tveksamt, välj det och tryck på Ctrl + Enter.
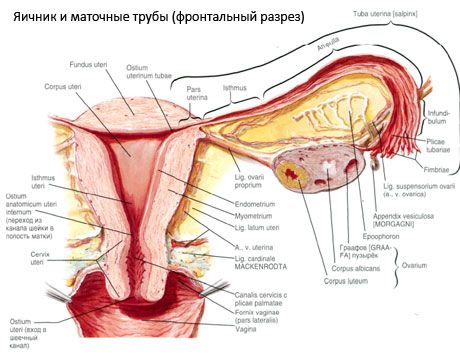
Ögon (äggstock, grekisk oophoron) - parat organ, kvinnligt könsorgan, lokaliserat i det lilla bäckens hål bakom livmoderns breda ligament. Äggstockar utvecklar och mognar kvinnliga könsceller (ägglossningar), liksom kvinnliga könshormoner som kommer in i blodet och lymfkörteln. Äggstocken har en ovoid form, något plattad i den främre och bakre riktningen. Färgen på äggstocken är rosa. På ytan av en kvinna som föds, är depression och ärr synliga - spår av ägglossning och omvandling av gula kroppar. äggstocks vikten var 8,5 g äggstocks dimensioner är :. Längd 2,5-5,5 cm, 1,5-3,0 cm bredd, tjocklek - upp till 2 cm har äggstocks skilja två fria ytor: en medial yta (facies medialis ) vänd mot bäckenhålan, delvis täckt äggledaren, och en lateral yta (facies later), intill den bäckensidoväggen, en mild fördjupning - äggstocks fossa. Denna fovea ligger i hörnet mellan de peritoneala yttre iliac kärlen på toppen och livmodern och ocklusala arterier i botten. Bakom äggstocken passerar urinledaren på motsvarande sida från toppen till botten.
Äggstocksytan rör sig i en konvex fri (bakre) kant (margo liber), framför - i den mesenteriala regionen (margo mesovaricus), med hjälp av ett kort klipp-faldigt av bukhinnan (tarmkäx äggstock) och baksidesbladet av breda ligament av livmodern. I förgrunden för kroppen är rafflade urtag - äggstock grind (hilum Ovarii), genom vilka äggstocken inkluderar artärer och nerver belägna vener och lymfkärl. Vi isolerade också äggstock två ändar: en rundad övre rörformig ände (extremitas tubaria), som vetter mot äggledaren, och den nedre änden av moderns (extremitas utenna), tillsammans med livmodern egen gäng äggstock (lig Ovarii proprium.). Denna bunt i en rundremmen runt 6 mm från modern är slutet till den laterala hörnet äggstocks uterin, som ligger mellan de två arken av breda ligament. Genom ligamentapparat äggstock också hänför ligament podveshivayaschaya äggstock (lig.suspensorium Ovarii), vilket är ett veck av bukhinnan som sträcker sig från den övre väggen av bäckenet till äggstocken och i äggstock innehållande kärl och fibrösa buntar av fibrer. Äggstock fast kort tarmkäxet (mesovarium), som representerar duplikatury bukhinnan, som sträcker sig från den bakre broschyren breda ligament av livmodern till äggstocken mesenteriska kanten. Äggstockarna själva täcks inte av bukhinnan. Den största äggstocksfibriaten i livmoderröret är fäst vid rörets ände av äggstocken. Ovariets topografi beror på livmoderns ställning, dess storlek (under graviditeten). Äggstockar hänvisar till mycket mobila organ i bäckenhålan.
 [
[Äggstockar och nerver
Blodtillförsel till äggstockarna beror på aa. Et vv. Ovaricae och uterinae. (. Aa ovaricae Dextra et sinistra) både äggstocksartären sträcker sig från den främre ytan av aorta just nedanför njurartärerna, rätten härstammar oftast från aortan och den vänstra - av njurartären. Att gå ner och i sidled över den främre ytan psoas muskler, skär äggstocks artär varje främre urinledare (ge honom kvistar), höftkärlen yttre och gränslinjen kommer in i bukhålan, som ligger här i den uppskjutande ligament i äggstockarna. Följande medialt, passerar äggstocksartären mellan arken av den breda ligament av livmodern under äggledarna, vilket ger grenar, och sedan - i tarmkäxet i äggstockarna; går in i äggstockarnas portar.
Blodkärlens grenar är i stor utsträckning anastomoserad med uterusartärens äggstocksgrenar. Venös utflöde från äggstockarna utförs främst i äggstocksvenös plexus, som ligger i äggstockarnas grind. Därför passerar utflödet av blod i två riktningar: genom livmodern och äggstockarna. Den högra äggstocksvenen har ventiler och går in i den nedre vena cava. Den vänstra äggstocksvenen strömmar in i vänster renalven utan några ventiler närvarande.
Lymfutflöde från äggstockarna sker genom lymfkärlen, särskilt rikligt i orginalportarna, där sublingualt lymfatisk plexus utsöndras. Därefter avleds lymfkörteln till de para-aorta lymfkörtlarna längs de ovarie-lymfatiska kärlen.
Innervation av äggstockarna
Sympatisk - tillhandahålls av postganglioniska fibrer från celiac (sol), övre flätad och hypogastrisk plexus; parasympatisk - på grund av de inre sakrala nerverna.
Struktur av äggstocken
Ytans yta är täckt med ett enkelskiktet germinalt epitel. Under den ligger en tät bind albuginea (tunica albuginea). Bindväv äggstocken former stroman (stroma Ovarii), rik på elastiska fibrer. Substans äggstocks parenkymet den delas upp av de yttre och inre skikten. Det inre skiktet, som ligger i centrum av äggstockarna, närmare porten, kallas medulla (medulla Ovarii). I detta skikt, i lös bindväv är många blod och lymfkärl och nerver. Yttre lager av äggstockar - cortex ovarii är mer tät. Det har en hel av bindväv, i vilken är belägna eftermogningskamrarna primära ovariefolliklar (folliculi ovarici primarii), sekundära (bubble) folliklar (folliculi ovarici secundarii, s.vesiculosi), en väl mogna folliklar graafovy bubblor (folliculi ovarici maturis), och gult och atretiska kroppar.
I varje follikel är en kvinnlig reproduktiv äggstock eller oocyt (ovocytus). Oocyt diameter upp till 150 mikron, rundade, innehåller en cellkärna, en stor mängd av cytoplasma, som förutom till cellulära organeller, det finns protein-lipid inneslutningar (äggula), glykogen behövs för ägg tillförsel. Dess oocyttillförsel förbrukar vanligen inom 12-24 timmar efter ägglossningen. Om befruktning inte uppstår, dör ägget.
Den mänskliga äggstocken har två täckmembran. Inuti är cytolemma, vilket är cytoklasmiska membranet hos oocyten. Utanför cytolemma finns det ett lager av så kallade follikelceller som skyddar ägget och uppfyller den hormonbildande funktionen - östrogenfrisättning.
Den fysiologiska positionen av livmodern, rören och äggstockarna tillhandahålls av suspensions-, fixerings- och stödapparaten, Kombination av peritoneum, ligament och bäckcellulosa. Suspensionsanordningen representeras av paraformationer, den innefattar runda och breda ledband i livmodern, egna ledband och hängande ligamentösa äggstockar. Livmoderns breda ligament, egen och upphängande ligament i äggstockarna håll livmodern i mittenpositionen. Runda ligament attraherar livmoderns botten främre och ger sin fysiologiska lutning.
Fastställande (fastställande) anordningen tillhandahåller en skakig position i centrum av en liten bassäng och gör det praktiskt taget omöjligt att kompensera delvis posteriort och anteriort. Men eftersom ligament i livmodern rör sig från sitt nedre delen, eventuell lutning av livmodern i olika riktningar. Av fixeringsenheten innefattar korder arrangerade i den lösa bäckenvävnaden och sträcker sig från den nedre livmoder kortet åt sidan, främre och bakre väggar av bäckenet: SI magochnye cardinal, livmoder och cystisk-Vesico-pubic ligamentet.
Förutom mesovarium utmärks följande äggstocksband:
- en suspension av äggstocksbandet, tidigare benämnt voronkotazovaya. Det är ett veck av bukhinnan med sträcker sig däri blod (a. Et v. Ovarica) och lymfatiska kärl och nerver ovarieceller sträckt mellan den laterala bäckenväggen, ryggradens fascia (i att dela den gemensamma höftartären vid den yttre och inre) och övre (rör) slutet av äggstocken;
- äggstockens egen ligament passerar mellan arken i det breda livmoderbandet, närmare det bakre arket, och förbinder den nedre änden av äggstocken med livmoderns sidomarginal. Till livmodern är äggstockens egen ligament fäst mellan början av livmoderröret och det cirkulära ligamentet, fram och tillbaka från sistnämnda. I tjockleken på ligamentet är rr. Ovarier, vilka är uterusartärens yttergrenar
- appendikulär-ovarie-ligamentet Klingan sträcker sig runt bilagans åsna till höger äggstocken eller livmoderns breda ligament i form av en vik i bukhinnan. Ledbandet är instabilt och observeras hos 1/2 - 1/3 av kvinnorna.
Stödapparaten representeras av bäckens golvets muskler och fasciae, uppdelad i nedre, mellersta och övre (inre) skikt.
Den mest kraftfulla är det övre (inre) muskelskiktet, representerat av den parade muskeln som ökar anusen. Den består av muskelknippen som fan ut från svanskotan till bäckenet i tre riktningar (blygd-svans, ilio-svans och ischiococcygeal muskler). Detta skikt av muskler kallas också bäckens membran.
Det mellersta lagret av muskler ligger mellan symfyserna, pubic och ischiumbenen. Mellersta lagret i musklerna - det urogenitala membranet - upptar den främre halvan av bäckens utlopp, genom det passerar urinröret och slidan. I mellan dess främre broschyrer är muskelknippen som bildar den yttre sfinktern i urinröret, i den bakre delen som muskelknippen som sträcker sig i en tvärriktning, - djupa tvärgående perineal muskel.
Den undre (yttre) skikt av bäckenbottenmuskulaturen består av ytan, vilken plats är formad som en figur 8. Dessa inkluderar bulbospongiosus-cavernous, ischio-cavernous, den yttre sfinktern av anus, den ytliga tvärgående perineal muskel.
Ontogeny av äggstockarna
Processen med tillväxt och follikulär atresi börjar med 20 veckors graviditet, och vid tidpunkten för leverans i flickornas äggstockar förblir upp till 2 miljoner oocyter. Vid tiden för menarche minskar deras antal till 300 tusen. Under hela perioden av reproduktiv liv når mognad och äggulerar högst 500 folliklar. Den initiala tillväxten av folliklar beror inte på stimulering av FSH, är begränsad, och atresi uppträder snabbt. Det antas att i stället för steroidhormoner är de lokala autokrin / parakrina peptiderna huvudregulatorn för tillväxt och atresi hos primära folliklar. Det antas att processen för tillväxt och atresi hos folliklarna inte avbryts av några fysiologiska processer. Denna process fortsätter i alla åldrar, inklusive intrauterin period och klimakteriet, avbryts av graviditet, ägglossning och anovulering. Mekanismen som utlöser tillväxten av folliklar och deras antal i varje specifik cykel är ännu inte klar.
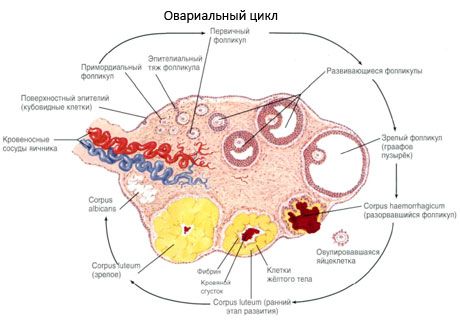
I sin utveckling genomgår follikeln flera utvecklingsstadier. Primordiala bakterieceller härrör från endosmen hos äggula sac, allanto och migrera till embryonets könsområde vid 5: e / 6: e graviditetsveckan. Som en följd av snabb mitotisk delning, som varar från 6-8 veckor till 16-20 veckors graviditet, bildas upp till 6-7 miljoner oocyter i embryonets äggstockar, omgivna av ett tunt lager av granulosa celler.
Den preantrala follikeln - oocyten är omgiven av ett membran (Zona pellucida). Granulosa celler som omger oocyten börjar prolifera, deras tillväxt beror på gonadotropinerna och korrelerar med östrogenhalten. Granulosa celler är målet för FSH. Vid stadium av preantral follikel kan granulosa celler syntetisera tre klasser av steroider: inducerar företrädesvis aktiviteten av aromatas, det huvudsakliga enzymet som omvandlar androgener till östradiol. Man tror att östradiol kan öka antalet egna receptorer, vilket ger en direkt mitogen effekt på granulosa celler oberoende av FSH. Det anses vara en parakrin faktor som förstärker effekterna av FSH, inklusive aktivering av aromatiseringsprocesser.
FSH-receptorer visas på membran av granulosa celler så snart tillväxten av follikeln börjar. Reduktion eller ökning av FSH leder till en förändring av antalet receptorer. Denna funktion av FSH moduleras av tillväxtfaktorer. FSH verkar via en G-protein adenylat-cyklassystemet i follikeln steroidogenes fastän huvudsakligen reglerad FSH, involverar processen många faktorer: jonkanaler, receptortyrosinkinas fosfolipas system av sekundära budbärare.
Androgenernas roll i follikelns tidiga utveckling är komplex. Granulosa-celler har androgenreceptorer. De är inte bara substrat för FSH-inducerad aromatisering till östrogen, men i låga koncentrationer kan förbättra processen för smakämnen. När nivån av androgener ökar preantral granulosaceller preferentiellt markerade banan ingen aromatisering till östrogen och enklare sätt att omvandling genom androgener 5a-reduktas i ett utvecklings androgen, som inte kan omvandlas till östrogen, och därmed inhiberade aromatasaktivitet. Denna process inhiberar också bildandet av FSH och LH-receptorer, och därigenom stoppa utvecklingen av follikeln.
Aromatiseringsprocessen, en follikel med en hög nivå av androgener, genomgår processer av atresi. Follikelns tillväxt och utveckling beror på dess förmåga att omvandla androgener till östrogener.
I närvaro av FSH är den dominerande substansen i follikelvätskan östrogener. I frånvaro av FSH - androgener. LH är normalt i follikelvätskan till mitten av cykeln. Så snart som den ökade nivån av mitotisk aktivitet hos LH granulosaceller minskar, och degenerativa förändringar verkar ökad androgen nivå i follikeln steroidnivåer i follikulär vätska än i plasma och återspeglar den funktionella aktiviteten äggstocken: granulosaceller och teka-celler. Om det enda målet för FSH granulosaceller, då LG har många mål - är teka celler, stromaceller och luteala och granulosaceller. Förmågan att steroidogenes har både granulosa och tekaceller, men aromatasaktiviteten dominerar i granulosa celler.
Som svar på LH producerar tekaceller androgener, vilka sedan genom FSH-inducerad aromatisering transformeras av granulosa celler till östrogener.
Som follikeln teka celler börjar uttrycka gener för LH-receptor P450 sek och 3beta-hydroxisteroiddehydrogenas, insulinliknande tillväxtfaktor (IGF-1) synergistiskt med LH för att öka uttrycket av genen, men inte stimulerar steroidogenes.
Ovarian steroidogenes är alltid LH-beroende. När follikeln växer uttrycker de nuvarande cellerna P450c17-enzymet, vilket bildar androgen från kolesterol. Granulosa celler har inte detta enzym och är beroende av de aktuella cellerna vid framställning av östrogener från androgener. Till skillnad från steroidogenes beror follikulogenes på FSH. Som follikeln och öka nivån av östrogen träder i funktion återkopplingsmekanism - inhiberade produktionen av FSH, vilket i sin tur leder till en minskning av aromatasaktivitet follikeln och, slutligen till follikeln atresi genom apoptos (programmerad celldöd).
Återkopplingsmekanismen för östrogener och FSH hämmar utvecklingen av folliklar som började växa, men inte den dominerande follikeln. Den dominerande follikeln innehåller mer FSH-receptorer som stöder proliferationen av granulosa celler och aromatiseringen av androgener i östrogener. Dessutom fungerar den parakrina och autokrina vägen som en viktig samordnare för utvecklingen av antralfollikeln.
En del autokrin / parakrin regulator är peptider (inhibin, aktivin, follistatin), granulosaceller som syntetiseras som svar på FSH och anländer i den follikulära vätskan. Inhibin reducerar FSH-sekretion; activin stimulerar frisättningen av FSH från hypofysen och ökar verkan av FSH i äggstocken; Follistatin undertrycker FSH-aktivitet, eventuellt på grund av bindning av aktivin. Efter ägglossning och utveckling av den gula kroppen är inhibin under kontroll av LH.
Tillväxten och differentieringen av äggstocksceller påverkas av insulinliknande tillväxtfaktorer (IGE). IGF-1 verkar på granulosa celler, vilket orsakar en ökning av cykliskt adenosinmonofosfat (cAMP), progesteron, oxytocin, proteoglykan och inhibin.
IGF-1 verkar på tekaceller vilket orsakar en ökning av produktionen av androgener. Tekakeller producerar i sin tur tumörnekrosfaktor (TNF) och epidermal tillväxtfaktor (EGF), som också regleras av FSH.
EGF stimulerar proliferation av granulosa celler. IGF-2 är huvudfaktorn för tillväxt av follikelvätska, detekterade också IGF-1, TNF-a, TNF-3 och EGF.
Brott mot parakrin och / eller autokrin reglering av ovariefunktionen verkar spela en roll vid störningar av ägglossningsprocesser och vid bildandet av polycystiska äggstockar.
När antralfollikeln växer ökar innehållet av östrogener i follikelvätskan. Vid toppen av deras ökning på granulosa celler uppträder receptorer för LH, luteinisering av granulosa celler uppträder och progesteronproduktionen ökar. Således under en preovulatorisk period orsakar en ökning av produktionen av östrogener utseendet av LH-receptorer, LH orsakar i sin tur luteinisering av granulosa-celler och progesteronproduktion. Ökningen av progesteron minskar nivån av östrogener, vilket tydligen orsakar den andra toppen av FSH i mitten av cykeln.
Det antas att ägglossningen sker 10-12 timmar efter toppen av LH och 24-36 timmar efter toppen av östradiol. Man tror att LH stimulerar reduktionen av oocyten, luteiniseringen av granulosa celler, syntesen av progesteron och prostaglandin i follikeln.
Progesteron ökar aktiviteten hos proteolytiska enzymer, tillsammans med prostaglandin involverat i att bryta follikelns vägg. FSH-inducerad progesteron topp, tillåter utsignalen från oocyten från follikeln genom att omvandla plasminogen till det proteolytiska enzymet - plasmin, ger en tillräcklig mängd av LH-receptorer för den normala utvecklingen av den luteala fasen.
Inom 3 dagar efter ägglossningen, granulosaceller ökar, verkar de karakteristiska vakuoler fyllda pigment - lutein. Teka-lutein-celler differentierar från teka och stroma och bli en del av corpus luteum. Mycket snabbt under inflytande av angiogena faktorer är utvecklingen av kapillärer som genomsyrar gulkroppen, och förbättring av kärl ökad produktion av progesteron och östrogen. Aktivitet steroidogenes och varaktighet corpus luteum liv bestäms av nivån på LH. Den gula kroppen är inte en homogen cellulär enhet. Tillsats av 2 typer luteal celler det innehåller endotelceller, makrofager, fibroblaster, och andra. Stora luteal celler producerar peptider (relaxin, oxytocin) och är mer aktiv i steroidogenes mer aromatasaktiviteten och en stor progesteronsyntes än de små cellerna.
Toppet av progesteron observeras den 8: e dagen efter toppen av LG. Det noterades att progesteron och östradiol i den luteala fasen utsöndras sporadiskt i samband med pulshastigheten hos LH. Med bildandet av en gul kropp kontrollerar produktionen av inhibinpass från FSH till LH. Ingibin ökar med ökningen av östradiol till toppen av LH och fortsätter att öka efter toppen av LH, även om östrogenhalten minskar. Fastän inhibin och östradiol utsöndras av granulosa celler, regleras de på olika sätt. Minskningen av inhibin vid slutet av lutealfasen bidrar till en ökning av FSH för nästa cykel.
Den gula kroppen mycket snabbt - på 9-11 dagen efter ägglossningen minskar.
Mekanismen för degenerering är inte tydlig och är inte relaterad till östrogenernas lyuteolitiska roll eller den receptorbundna mekanismen, vilket ses i endometrium. Det finns en annan förklaring till rollen av östrogener som produceras av den gula kroppen. Det är känt att för syntes av progesteronreceptorer i endometrium krävs östrogener. Lutealfas östrogener är förmodligen nödvändiga för progesteronrelaterade förändringar i endometrium efter ägglossningen. Otillräcklig utveckling av progesteronreceptorerna, som ett resultat av otillräckligt östrogeninnehåll, är förmodligen en ytterligare mekanism för infertilitet och tidig graviditetsförlust, en annan form av underlägsen lutealfas. Man tror att livslängden hos den gula kroppen är inställd vid tidpunkten för ägglossningen. Och det kommer säkert att regresseras om choriongonadotropin inte stöds i samband med graviditet. Således leder regression av den gula kroppen till en minskning av nivåerna av östradiol, progesteron och inhibin. Reduktionshämman avlägsnar dess hämmande effekt på FSH; reduktion av östradiol och progesteron möjliggör mycket snabbt att återställa sekretionen av GnRH och avlägsna mekanismen för återkoppling från hypofysen. Reduktion av inhibin och östradiol, tillsammans med en ökning av GnRH, leder till prevalens av FSH över LH. En ökning av FSH leder till tillväxten av folliklar med det efterföljande valet av den dominerande follikeln, och en ny cykel börjar, om en graviditet inte uppstår. Steroidhormoner spelar en ledande roll inom reproduktiv biologi och i allmänhet fysiologi. De bestämmer fenotypen hos en person, påverkar hjärt-kärlsystemet, ämnesomsättningen av ben, hud, kroppens allmänna välbefinnande och spelar en nyckelroll vid graviditeten. Verkan av steroidhormoner återspeglar de intracellulära och genetiska mekanismerna som är nödvändiga för att överföra den extracellulära signalen till cellens kärna för att inducera ett fysiologiskt svar.
Estrogener diffunderar genom cellmembranet och binder till receptorer belägna i cellens kärna. Receptor-steroidkomplexet bindar sedan till DNA. I målceller leder dessa interaktioner till genuttryck, syntesen av proteiner, till en specifik funktion av celler och vävnader.