Medicinsk expert av artikeln

Nya publikationer
hypofysen
Senast recenserade: 04.07.2025

Allt iLive-innehåll är mediekontrollerat eller faktiskt kontrollerat för att säkerställa så mycket faktuell noggrannhet som möjligt.
Vi har strikta sourcing riktlinjer och endast länk till välrenommerade media webbplatser, akademiska forskningsinstitut och, när det är möjligt, medicinsk peer granskad studier. Observera att siffrorna inom parentes ([1], [2] etc.) är klickbara länkar till dessa studier.
Om du anser att något av vårt innehåll är felaktigt, omodernt eller på annat sätt tveksamt, välj det och tryck på Ctrl + Enter.
Hypofysen (hypofysen, s.glandula pituitaria) är belägen i hypofysens fossa i kilbenet (sella turcica) och är separerad från kranialhålan genom en utskjutande del av dura mater i hjärnan, som bildar sellas diafragma. Genom öppningen i detta diafragma är hypofysen ansluten till infundibulum i hypotalamus i diencephalon. Hypofysens tvärgående storlek är 10-17 mm, anteroposterior är 5-15 mm, vertikal är 5-10 mm. Hypofysens massa hos män är cirka 0,5 g, hos kvinnor - 0,6 g. Hypofysen är täckt utvändigt av en kapsel.
I enlighet med hypofysens utveckling från två olika grundämnen, urskiljs två lober i organet - den främre och den bakre. Adenohypofysen, eller den främre loben (adenohypophysis, s.lobus anterior), är större och utgör 70-80% av hypofysens totala massa. Den är tätare än den bakre loben. I den främre loben urskiljs en distal del (pars distalis), som upptar den främre delen av hypofysgropen, en mellanliggande del (pars intermedia), som är belägen på gränsen till den bakre loben, och en knölformad del (pars tuberalis), som går uppåt och ansluter till hypotalamus infundibulum. På grund av överflödet av blodkärl har den främre loben en ljusgul färg med en rödaktig nyans. Parenkym i den främre hypofysen representeras av flera typer av körtelceller, mellan vilkas strängar sinusformade blodkapillärer är belägna. Hälften (50 %) av adenohypofyscellerna är kromofila adenocyter, som har finkorniga granuler i sin cytoplasma som färgas väl med kromsalter. Dessa är acidofila adenocyter (40 % av alla adenohypofysceller) och basofila adenocyter (10 %). Basofila adenocyter inkluderar gonadotropa, kortikotropa och tyreotropa endokrinocyter. Kromofoba adenocyter är små, de har en stor kärna och en liten mängd cytoplasma. Dessa celler anses vara föregångare till kromofila adenocyter. De övriga 50 % av adenohypofyscellerna är kromofoba adenocyter.
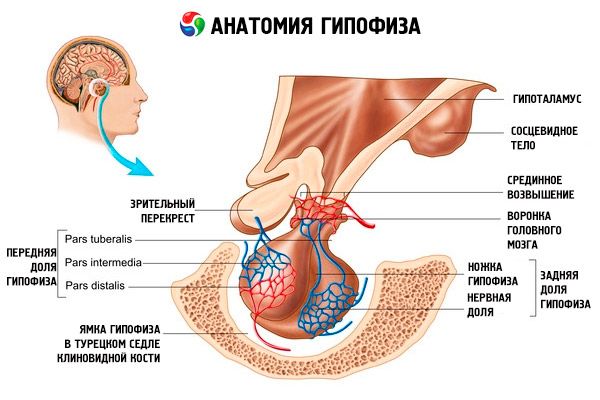
Neurohypofysen, eller bakre loben (neurohypophysis, s.lobus posterior), består av nervloben (lobus nervosus), som är belägen i den bakre delen av hypofysens fossa, och tratten (infundibulum), som är belägen bakom den tuberösa delen av adenohypofysen. Hypofysens bakre lob bildas av neurogliaceller (hypofysceller), nervfibrer som går från hypotalamus neurosekretoriska kärnor till neurohypofysen, och neurosekretoriska blodkroppar.
Hypofysen är, via nervfibrer (banor) och blodkärl, funktionellt kopplad till hypotalamus i diencephalon, som reglerar hypofysens aktivitet. Hypofysen och hypotalamus, tillsammans med deras neuroendokrina, kärl- och nervförbindelser, betraktas vanligtvis som hypotalamus-hypofyssystemet.
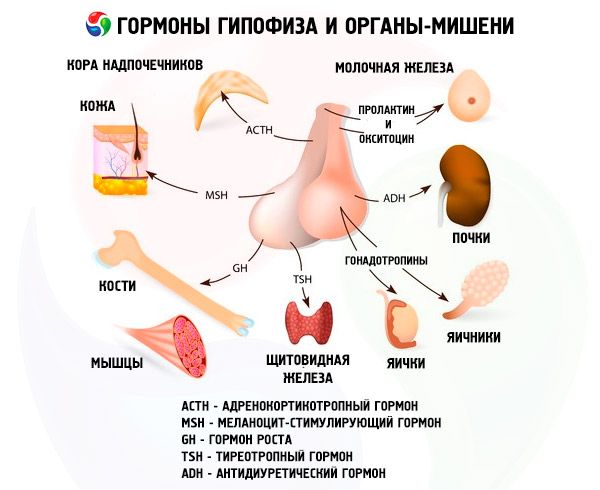
Hormoner i främre och bakre hypofysen påverkar många av kroppens funktioner, främst genom andra endokrina körtlar. I den främre hypofysen producerar acidofila adenocyter (alfaceller) somatotropiskt hormon (tillväxthormon), vilket deltar i regleringen av tillväxt- och utvecklingsprocesser hos unga organismer. Kortikotropa endokrinocyter utsöndrar adrenokortikotropiskt hormon (ACTH), vilket stimulerar binjurarnas utsöndring av steroidhormoner. Tyreotropa endokrinocyter utsöndrar tyreotropiskt hormon (TSH), vilket påverkar sköldkörtelns utveckling och aktiverar produktionen av dess hormoner. Gonadotropa hormoner: follikelstimulerande (FSH), luteiniserande (LH) och prolaktin - påverkar kroppens sexuella mognad, reglerar och stimulerar utvecklingen av folliklar i äggstockarna, ägglossning, tillväxt av mjölkkörtlarna och mjölkproduktion hos kvinnor, samt spermatogenesprocessen hos män. Dessa hormoner produceras av basofila adenocyter (betaceller). Lipotropa faktorer från hypofysen utsöndras också här, vilka påverkar mobiliseringen och utnyttjandet av fetter i kroppen. I den mellanliggande delen av den främre loben bildas melanocytstimulerande hormon, vilket styr bildandet av pigment - melaniner - i kroppen.
Neurosekretoriska celler i de supraoptiska och paraventrikulära kärnorna i hypotalamus producerar vasopressin och oxytocin. Dessa hormoner transporteras till cellerna i den bakre hypofysen längs axoner som utgör hypotalamus-hypofysen. Från den bakre hypofysen kommer dessa ämnen in i blodomloppet. Hormonet vasopressin har en vasokonstriktiv och antidiuretisk effekt, vilket det också kallas antidiuretiskt hormon (ADH). Oxytocin har en stimulerande effekt på livmodermusklernas kontraktilitet, ökar mjölksekretionen från den ammande mjölkkörteln, hämmar utvecklingen och funktionen av corpus luteum och påverkar förändringar i tonusen i de glatta (icke-strimmiga) musklerna i mag-tarmkanalen.
Utveckling av hypofysen
Hypofysens främre lob utvecklas från epitelet i munhålans dorsala vägg i form av en ringformad utväxt (Rathkes påse). Denna ektodermala utväxt växer mot botten av den framtida tredje ventrikeln. Mot den, från den nedre ytan av den andra hjärnblåsan (den framtida botten av den tredje ventrikeln), växer en process, från vilken den grå tuberkeln i infundibulum och hypofysens bakre lob utvecklas.
Kärl och nerver i hypofysen
Från de inre halspulsådrorna och kärlen i storhjärnans artärcirkel leds de övre och nedre hypofysartärerna till hypofysen. De övre hypofysartärerna går till den grå kärnan och hypotalamus infundibulum, anastomoserar med varandra här och bildar kapillärer som penetrerar hjärnvävnaden - det primära hemokapillära nätverket. Från de långa och korta slingorna i detta nätverk bildas portvenerna, vilka leds till hypofysens främre lob. I parenkymet i hypofysens främre lob bryts dessa vener upp i breda sinusformade kapillärer och bildar det sekundära hemokapillära nätverket. Hypofysens bakre lob försörjs huvudsakligen med blod från den nedre hypofysartären. Mellan de övre och nedre hypofysartärerna finns långa arteriella anastomoser. Utflödet av venöst blod från det sekundära hemokapillära nätverket sker genom ett system av vener som rinner in i de kavernösa och interkavernösa bihålorna i hjärnans dura mater.
Sympatiska fibrer som kommer in i organet tillsammans med artärer deltar i hypofysens innervation. Postganglionära sympatiska nervfibrer utgår från plexus i den inre halspulsådern. Dessutom finns många ändelser av processerna hos neurosekretoriska celler, belägna i hypotalamuskärnan, i hypofysens bakre lob.
Åldersrelaterade egenskaper hos hypofysen
Hypofysens genomsnittliga vikt hos nyfödda når 0,12 g. Organets vikt fördubblas vid 10 års ålder och tredubblas vid 15 års ålder. Vid 20 års ålder når hypofysens vikt sitt maximum (530-560 mg) och förblir nästan oförändrad under efterföljande åldersperioder. Efter 60 år observeras en liten minskning av vikten av denna endokrina körtel.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hypofyshormoner
Enheten i nervös och hormonell reglering i kroppen säkerställs av den nära anatomiska och funktionella kopplingen mellan hypofysen och hypotalamus. Detta komplex bestämmer tillståndet och funktionen hos hela det endokrina systemet.
Den huvudsakliga endokrina körteln som producerar ett antal peptidhormoner som direkt reglerar funktionen hos de perifera körtlarna är hypofysen. Det är en rödgrå bönformad formation täckt med en fibrös kapsel som väger 0,5-0,6 g. Den varierar något beroende på kön och ålder. Den allmänt accepterade uppdelningen av hypofysen i två lober som skiljer sig åt i utveckling, struktur och funktion kvarstår: den främre distala - adenohypofysen och den bakre - neurohypofysen. Den första utgör cirka 70% av körtelns totala massa och är konventionellt uppdelad i den distala, infundibulära och mellanliggande delen, den andra - i den bakre delen, eller loben, och hypofysens stjälk. Körteln är belägen i hypofysens fossa i sella turcica i kilbenet och är ansluten till hjärnan genom stjälken. Den övre delen av den främre loben är täckt av synkiasmen och synkanalerna. Blodtillförseln till hypofysen är mycket riklig och sker via grenar av den inre halspulsådern (hypofysartärerna övre och nedre), samt via grenar av hjärnans artärcirkel. De övre hypofysartärerna deltar i blodtillförseln till adenohypofysen, och de nedre - neurohypofysen, och kommer i kontakt med de neurosekretoriska ändarna på axonerna i hypotalamus stora cellkärnor. De förra når hypotalamus mediana eminens, där de sprider sig i ett kapillärnätverk (primär kapillärplexus). Dessa kapillärer (med vilka axonterminalerna i de små neurosekretoriska cellerna i den mediobasala hypotalamus kommer i kontakt) samlas i portalvener som går ner längs hypofysstjälken in i adenohypofysens parenkym, där de återigen delar sig i ett nätverk av sinusformade kapillärer (sekundär kapillärplexus). Således kommer blodet, som tidigare passerat genom hypotalamus median eminens, där det berikas med hypotalamiska adenohypofysotropa hormoner (frisättande hormoner), in i adenohypofysen.
Utflödet av blod mättat med adenohypofyshormoner från många kapillärer i sekundärplexus sker genom vensystemet, som i sin tur strömmar in i dura maters venösa bihålor och sedan in i den allmänna blodomloppet. Således är hypofysens portalsystem med en nedåtgående riktning för blodflödet från hypotalamus en morfofunktionell komponent i den komplexa mekanismen för neurohumoral kontroll av adenohypofysens tropiska funktioner.
Hypofysen innerveras av sympatiska fibrer som följer hypofysartärerna. De utgår från postganglionära fibrer som passerar genom den interna carotisplexusen, anslutna till de övre cervikala ganglierna. Det finns ingen direkt innervation av adenohypofysen från hypotalamus. Den bakre loben tar emot nervfibrer från hypotalamus neurosekretoriska kärnor.
Adenohypofysen är en mycket komplex formation i sin histologiska arkitektur. Den har två typer av körtelceller - kromofoba och kromofila. De senare är i sin tur indelade i acidofila och basofila (en detaljerad histologisk beskrivning av hypofysen ges i motsvarande avsnitt i manualen). Det bör dock noteras att hormonerna som produceras av körtelcellerna som utgör adenohypofysens parenkym, på grund av de senares mångfald, till viss del skiljer sig åt i sin kemiska natur, och den fina strukturen hos de utsöndrande cellerna måste motsvara biosyntesegenskaperna hos var och en av dem. Men ibland kan övergångsformer av körtelceller som kan producera flera hormoner observeras i adenohypofysen. Det finns bevis för att typen av körtelceller i adenohypofysen inte alltid är genetiskt bestämd.
Under diafragman på sella turcica sitter den trattformade delen av den främre loben. Den omsluter hypofysens stjälk och är i kontakt med den grå tuberkeln. Denna del av adenohypofysen kännetecknas av närvaron av epitelceller och riklig blodtillförsel. Den är också hormonellt aktiv.
Den mellanliggande (mellersta) delen av hypofysen består av flera lager av stora sekretoriskt aktiva basofila celler.
Hypofysen utför olika funktioner genom sina hormoner. Dess främre lob producerar adrenokortikotropa (ACTH), tyreoidstimulerande (TSH), follikelstimulerande (FSH), luteiniserande (LH), lipotropa hormoner, samt tillväxthormon - somatotropiskt (STO) och prolaktin. I den mellanliggande loben syntetiseras melanocytstimulerande hormon (MSH), och i den bakre loben ackumuleras vasopressin och oxytocin.
ACTH
Hypofyshormoner är en grupp av protein- och peptidhormoner och glykoproteiner. Av hormonerna i den främre hypofysen har ACTH studerats bäst. Det produceras av basofila celler. Dess huvudsakliga fysiologiska funktion är att stimulera biosyntesen och utsöndringen av steroidhormoner från binjurebarken. ACTH uppvisar också melanocytstimulerande och lipotropisk aktivitet. År 1953 isolerades det i ren form. Senare fastställdes dess kemiska struktur, bestående av 39 aminosyrarester hos människor och ett antal däggdjur. ACTH har ingen artspecificitet. För närvarande har kemisk syntes av både själva hormonet och olika fragment av dess molekyl, mer aktiva än naturliga hormoner, genomförts. Hormonets struktur har två sektioner av peptidkedjan, varav den ena säkerställer detektion och bindning av ACTH till receptorn, och den andra ger en biologisk effekt. Det binder tydligen till ACTH-receptorn genom interaktionen mellan hormonets och receptorns elektriska laddningar. Rollen som den biologiska effektorn för ACTH utförs av fragmentet av 4-10-molekylen (Met-Glu-His-Phen-Arg-Tri-Tri).
ACTHs melanocytstimulerande aktivitet beror på närvaron i molekylen av den N-terminala regionen, som består av 13 aminosyrarester och upprepar strukturen hos alfa-melanocytstimulerande hormon. Denna region innehåller också en heptapeptid som finns i andra hypofyshormoner och som har viss adrenokortikotropisk, melanocytstimulerande och lipotropisk aktivitet.
Det avgörande momentet i ACTH:s verkan anses vara aktiveringen av enzymet proteinkinas i cytoplasman med deltagande av cAMP. Fosforylerat proteinkinas aktiverar enzymet esteras, vilket omvandlar kolesterolestrar till en fri substans i fettdropparna. Proteinet som syntetiseras i cytoplasman som ett resultat av fosforylering av ribosomer stimulerar bindningen av fritt kolesterol till cytokrom P-450 och dess överföring från lipiddroppar till mitokondrier, där alla enzymer som säkerställer omvandlingen av kolesterol till kortikosteroider finns.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Sköldkörtelstimulerande hormon
TSH - tyreotropin - den huvudsakliga regulatorn för sköldkörtelns utveckling och funktion, samt processerna för syntes och utsöndring av sköldkörtelhormoner. Detta komplexa protein - glykoprotein - består av alfa- och beta-subenheter. Strukturen för den första subenheten sammanfaller med alfa-subenheten för luteiniserande hormon. Dessutom sammanfaller den i stort sett hos olika djurarter. Sekvensen av aminosyrarester i beta-subenheten för mänsklig TSH har dechiffrerats och består av 119 aminosyrarester. Det kan noteras att beta-subenheterna i mänsklig och nötkreaturs TSH är likartade på många sätt. De biologiska egenskaperna och naturen hos glykoproteinhormonernas biologiska aktivitet bestäms av beta-subenheten. Den säkerställer också hormonets interaktion med receptorer i olika målorgan. Beta-subenheten uppvisar dock specifik aktivitet hos de flesta djur först efter att den kombinerats med alfa-subenheten, som fungerar som en slags aktivator för hormonet. Den senare inducerar med lika stor sannolikhet luteiniserande, follikelstimulerande och tyreotropa aktiviteter som bestäms av beta-subenhetens egenskaper. Den funna likheten gör att vi kan dra slutsatsen att dessa hormoner uppstod i evolutionsprocessen från en gemensam föregångare, beta-subenheten bestämmer också hormonernas immunologiska egenskaper. Det finns ett antagande att alfa-subenheten skyddar beta-subenheten från proteolytiska enzymers verkan och även underlättar dess transport från hypofysen till de perifera målorganen.
Gonadotropa hormoner
Gonadotropiner finns i kroppen i form av LH och FSH. Det funktionella syftet med dessa hormoner är generellt reducerat till att säkerställa reproduktionsprocesser hos individer av båda könen. De är, liksom TSH, komplexa proteiner - glykoproteiner. FSH inducerar mognad av folliklar i kvinnors äggstockar och stimulerar spermatogenesen hos män. LH orsakar follikelruptur hos kvinnor med bildandet av corpus luteum och stimulerar utsöndringen av östrogener och progesteron. Hos män accelererar samma hormon utvecklingen av interstitiell vävnad och utsöndringen av androgener. Effekterna av gonadotropiner är beroende av varandra och sker synkront.
Dynamiken i gonadotropinsekretionen hos kvinnor förändras under menstruationscykeln och har studerats tillräckligt noggrant. I den preovulatoriska (follikulära) fasen av cykeln är LH-halten på en ganska låg nivå, medan FSH ökar. När follikeln mognar ökar östradiolsekretionen, vilket främjar ökad produktion av gonadotropiner från hypofysen och förekomsten av både LH- och FSH-cykler, dvs. könshormoner stimulerar utsöndringen av gonadotropiner.
För närvarande har strukturen för LH fastställts. Liksom TSH består den av två subenheter: a och b. Strukturen för alfa-subenheten i LH hos olika djurarter överensstämmer i stort sett med strukturen för alfa-subenheten i TSH.
Strukturen för beta-subenheten i LH skiljer sig avsevärt från strukturen för beta-subenheten i TSH, även om den har fyra identiska sektioner av peptidkedjan bestående av 4-5 aminosyrarester. I TSH är de lokaliserade i positionerna 27-31, 51-54, 65-68 och 78-83. Eftersom beta-subenheten i LH och TSH bestämmer hormonernas specifika biologiska aktivitet, kan man anta att homologa sektioner i strukturen hos LH och TSH bör säkerställa kopplingen mellan beta-subenheterna och alfa-subenheten, och sektioner som skiljer sig åt i struktur bör vara ansvariga för specificiteten hos hormonernas biologiska aktivitet.
Naturligt LH är mycket stabilt mot proteolytiska enzymers verkan, men beta-subenheten klyvs snabbt av kymotrypsin, och a-subenheten är svår att hydrolysera av enzymet, dvs. den spelar en skyddande roll och förhindrar kymotrypsin från att komma åt peptidbindningarna.
När det gäller den kemiska strukturen hos FSH har forskare ännu inte fått fram definitiva resultat. Liksom LH består FSH av två subenheter, men beta-subenheten hos FSH skiljer sig från beta-subenheten hos LH.
Prolaktin
Ett annat hormon, prolaktin (laktogent hormon), spelar en aktiv roll i reproduktionsprocesserna. De viktigaste fysiologiska egenskaperna hos prolaktin hos däggdjur manifesteras i form av stimulering av utvecklingen av mjölkkörtlar och laktation, tillväxt av talgkörtlar och inre organ. Det främjar manifestationen av steroiders effekt på sekundära sexuella egenskaper hos hanar, stimulerar den gulkroppens sekretoriska aktivitet hos möss och råttor och deltar i regleringen av fettmetabolismen. Mycket uppmärksamhet har ägnats åt prolaktin under senare år som en regulator av moderns beteende; sådan polyfunktionalitet förklaras av dess evolutionära utveckling. Det är ett av de gamla hypofyshormonerna och finns även hos amfibier. För närvarande har prolaktins struktur hos vissa däggdjursarter blivit helt dechiffrerad. Men fram till nyligen uttryckte forskare tvivel om förekomsten av ett sådant hormon hos människor. Många trodde att dess funktion utfördes av tillväxthormonet. Nu har övertygande bevis erhållits för förekomsten av prolaktin hos människor och dess struktur har delvis dechiffrerats. Prolaktinreceptorer binder aktivt tillväxthormon och placentalaktogen, vilket indikerar en enda verkningsmekanism för de tre hormonerna.
Somatotropin
Tillväxthormonet somatotropin har ett ännu bredare verkningsspektrum än prolaktin. Liksom prolaktin produceras det av acidofila celler i adenohypofysen. Somatotropin stimulerar skeletttillväxt, aktiverar proteinbiosyntes, har en fettmobiliserande effekt och främjar en ökning av kroppsstorlek. Dessutom koordinerar det metaboliska processer.
Hormonets deltagande i det senare bekräftas av en kraftig ökning av dess utsöndring av hypofysen, till exempel när blodsockernivån sjunker.
Den kemiska strukturen för detta mänskliga hormon är nu helt fastställd - 191 aminosyrarester. Dess primära struktur liknar den för korioniskt somatomammotropin eller placentalaktogen. Dessa data indikerar en betydande evolutionär närhet mellan de två hormonerna, även om de uppvisar skillnader i biologisk aktivitet.
Det är nödvändigt att betona den höga artspecificiteten hos det aktuella hormonet - till exempel är STH av animaliskt ursprung inaktivt hos människor. Detta förklaras av både reaktionen mellan STH-receptorerna hos människor och djur, och hormonets struktur i sig. För närvarande pågår studier för att identifiera aktiva centra i STH:s komplexa struktur som uppvisar biologisk aktivitet. Enskilda fragment av molekylen som uppvisar andra egenskaper studeras. Till exempel, efter hydrolys av humant STH med pepsin, isolerades en peptid bestående av 14 aminosyrarester och motsvarande molekylavsnitt 31-44. Den hade ingen tillväxteffekt, men överträffade signifikant det nativa hormonet i lipotropisk aktivitet. Humant tillväxthormon, till skillnad från liknande hormoner hos djur, har betydande laktogen aktivitet.
Adenohypofysen syntetiserar många peptid- och proteinsubstanser som har en fettmobiliserande effekt, och de hypofysiska tropikhormonerna - ACTH, STH, TSH och andra - har en lipotropisk effekt. På senare år har beta- och y-lipotropiska hormoner (LPG) särskilt betonats. De biologiska egenskaperna hos beta-LPG har studerats mycket detaljerat; förutom lipotropisk aktivitet har det också en melanocytstimulerande, kortikotropinstimulerande och hypokalcemisk effekt, och ger även en insulinliknande effekt.
För närvarande har den primära strukturen för får-LPG (90 aminosyrarester), lipotropa hormoner hos grisar och nötkreatur, dechiffrerats. Detta hormon har artspecificitet, även om strukturen i den centrala regionen av beta-LPG är densamma hos olika arter. Det bestämmer hormonets biologiska egenskaper. Ett av fragmenten av denna region finns i strukturen hos alfa-MSH, beta-MSH, ACTH och beta-LPG. Det föreslås att dessa hormoner uppstod från samma prekursor i evolutionsprocessen. γ-LPG har en svagare lipotropisk aktivitet än beta-LPG.
Melanocytstimulerande hormon
Detta hormon, som syntetiseras i hypofysens mellanlob, stimulerar biosyntesen av hudpigmentet melanin i dess biologiska funktion, främjar en ökning av storleken och antalet pigmentceller melanocyter i amfibiers hud. Dessa egenskaper hos MSH används i biologiska tester av hormonet. Det finns två typer av hormonet: alfa- och beta-MSH. Det har visats att alfa-MSH inte har artspecificitet och har samma kemiska struktur hos alla däggdjur. Dess molekyl är en peptidkedja bestående av 13 aminosyrarester. Beta-MSH har däremot artspecificitet, och dess struktur skiljer sig åt hos olika djur. Hos de flesta däggdjur består beta-MSH-molekylen av 18 aminosyrarester, och endast hos människor är den förlängd från aminosyraänden med fyra aminosyrarester. Det bör noteras att alfa-MSH har viss adrenokortikotropisk aktivitet, och dess effekt på djurs och människors beteende har nu bevisats.
Oxytocin och vasopressin
Vasopressin och oxytocin, som syntetiseras i hypotalamus, ackumuleras i hypofysens bakre lob: vasopressin i neuronerna i den supraoptiska kärnan och oxytocin i den paraventrikulära kärnan. Därefter överförs de till hypofysen. Det bör betonas att föregångaren till vasopressinhormonet syntetiseras först i hypotalamus. Samtidigt produceras proteinet neurofysin typ 1 och 2 där. Det första binder oxytocin, och det andra binder vasopressin. Dessa komplex migrerar i form av neurosekretoriska granuler i cytoplasman längs axonet och når hypofysens bakre lob, där nervfibrerna slutar i kärlväggen och granulernas innehåll kommer in i blodomloppet. Vasopressin och oxytocin är de första hypofyshormonerna med en fullständigt etablerad aminosyrasekvens. I sin kemiska struktur är de nonapeptider med en disulfidbrygga.
De hormoner som behandlas har olika biologiska effekter: de stimulerar transporten av vatten och salter genom membran, har en vasopressorisk effekt, förstärker sammandragningar av livmoderns glatta muskulatur under förlossningen och ökar utsöndringen av mjölkkörtlarna. Det bör noteras att vasopressin har en högre antidiuretisk aktivitet än oxytocin, medan det senare har en starkare effekt på livmodern och mjölkkörtlarna. Den huvudsakliga regulatorn av vasopressinsekretion är vattenintag; i njurtubuli binder det till receptorer i de cytoplasmatiska membranen med efterföljande aktivering av enzymet adenylatcyklas i dem. Olika delar av molekylen ansvarar för att binda hormonet till receptorn och för den biologiska effekten.
Hypofysen, som är förbunden via hypotalamus med hela nervsystemet, förenar det endokrina systemet till en funktionell helhet och bidrar till att säkerställa kroppens inre miljö (homeostas). Inom det endokrina systemet sker homeostatisk reglering baserat på principen om återkoppling mellan hypofysens främre lob och "målkörtlarna" (sköldkörteln, binjurebarken, gonader). Överskott av hormonet som produceras av "målkörteln" hämmar, och dess brist stimulerar, utsöndringen och frisättningen av motsvarande tropiska hormon. Hypotalamus ingår i återkopplingssystemet. Det är i det som receptorzonerna som är känsliga för hormonerna i "målkörtlarna" finns. Genom att specifikt binda till hormoner som cirkulerar i blodet och ändra responsen beroende på hormonkoncentrationen, överför hypotalamiska receptorer sin effekt till motsvarande hypotalamiska centra, som koordinerar adenohypofysens arbete och frisätter hypotalamiska adenohypofysiotropa hormoner. Således bör hypotalamus betraktas som en neuroendokrin hjärna.