Medicinsk expert av artikeln

Nya publikationer
Kolera vibrio
Senast recenserade: 04.07.2025

Allt iLive-innehåll är mediekontrollerat eller faktiskt kontrollerat för att säkerställa så mycket faktuell noggrannhet som möjligt.
Vi har strikta sourcing riktlinjer och endast länk till välrenommerade media webbplatser, akademiska forskningsinstitut och, när det är möjligt, medicinsk peer granskad studier. Observera att siffrorna inom parentes ([1], [2] etc.) är klickbara länkar till dessa studier.
Om du anser att något av vårt innehåll är felaktigt, omodernt eller på annat sätt tveksamt, välj det och tryck på Ctrl + Enter.
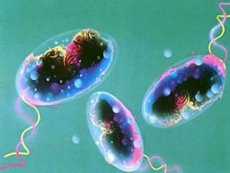
Enligt WHO är kolera en infektionssjukdom som kännetecknas av akut, svår, uttorkande diarré med avföring i form av risvatten, vilket är en följd av infektion med Vibrio cholerae. På grund av att den kännetecknas av en uttalad förmåga att sprida sig vid epidemier, ett svårt förlopp och en hög dödlighet, anses kolera vara en särskilt farlig infektion.
Kolerans historiska hemland är Indien, eller mer exakt, deltat i floderna Ganges och Brahmaputra (nuvarande Östindien och Bangladesh), där den har funnits sedan urminnes tider (koleraepidemier i denna region observerades så tidigt som 500 f.Kr.). Den långa existensen av ett endemiskt fokus för kolera här förklaras av många anledningar. Koleravibrion kan inte bara överleva i vatten under lång tid, utan också föröka sig i det under gynnsamma förhållanden - temperaturer över 12 °C, närvaron av organiskt material. Alla dessa förhållanden är tydliga i Indien: ett tropiskt klimat (genomsnittlig årstemperatur från 25 till 29 °C), riklig nederbörd och träskmark, hög befolkningstäthet, särskilt i Gangesflodens delta, en stor mängd organiskt material i vattnet, kontinuerlig vattenförorening med avloppsvatten och exkrementer året runt, en låg materiell levnadsstandard och unika religiösa och kultistiska riter hos befolkningen.
I koleraepidemiernas historia kan fyra perioder urskiljas.
Period I - fram till 1817, då kolera endast var koncentrerad i Öst- och Sydasien, främst i Indien, och inte spred sig utanför dess gränser.
II-perioden - från 1817 till 1926. Med etablerandet av breda ekonomiska och andra band mellan Indien och europeiska och andra länder, spred sig kolera bortom Indien och orsakade, genom att spreda sig längs ekonomiska och religiösa band, 6 pandemier som krävde miljontals människoliv. Ryssland var det första av de europeiska länderna där kolera spred sig. Från 1823 till 1926 upplevde Ryssland 57 koleraår. Under denna tid insjuknade mer än 5,6 miljoner människor i kolera och 2,14 miljoner människor dog av den ("40%).
III-perioden - från 1926 till 1961 återgick koleran till sitt huvudsakliga endemiska fokus, och en period av relativt välbefinnande började. Det verkade som att med utvecklingen av moderna system för rening av dricksvatten, avlägsnande och desinfektion av avloppsvatten och utveckling av speciella åtgärder mot kolera, inklusive skapandet av en karantäntjänst, skulle världens länder vara tillförlitligt skyddade från ytterligare en kolerainvasion.
Den fjärde perioden började 1961 och fortsätter än idag. Den sjunde pandemin började inte i Indien, utan i Indonesien, spred sig snabbt till Filippinerna, Kina, Indokina och sedan till andra länder i Asien, Afrika och Europa. Särdragen hos denna pandemi inkluderar det faktum att den för det första orsakades av en speciell variant av koleravibrion - V. cholerae eltor, som fram till 1961 inte ens officiellt erkändes som orsaken till kolera; för det andra, vad gäller varaktighet, överträffade den alla tidigare pandemier; för det tredje inträffade den i två vågor, varav den första varade till 1990, och den andra började 1991 och omfattade många länder i Syd- och Nordamerika, inklusive USA, som inte hade sett en koleraepidemi sedan 1866. Från 1961 till 1996 insjuknade 3 943 239 personer i kolera i 146 länder.
Koleras orsakande agens, Vibrio cholerae, upptäcktes 1883 under den femte pandemin av R. Koch, men vibrio upptäcktes först i avföringen från patienter med diarré 1854 av F. Pacini.
V. cholerae tillhör familjen Vibrionaceae, som omfattar flera släkten (Vibrio, Aeromonas, Plesiomonas, Photobacterium). Släktet Vibrio har mer än 25 arter sedan 1985, varav de viktigaste för människor är V. cholerae, V. parahaemolyticus, V. alginolyticus, V. vulnificus och V. fluvialis.
Viktiga egenskaper hos släktet Vibrio
Korta, icke-spor- och kapselbildande, böjda eller raka gramnegativa stavar, 0,5 µm i diameter och 1,5-3,0 µm långa, rörliga (V. cholerae är monotrik, vissa arter har två eller flera polära flageller); växer bra och snabbt på vanliga medier, är kemoorganotrofer och fermenterar kolhydrater för att producera syra utan gas (glukos fermenteras via Embden-Meyerhof-vägen). Oxidaspositiva, bildar indol, reducerar nitrater till nitriter (V. cholerae ger en positiv nitrosoindolreaktion), bryter ner gelatin, ger ofta en positiv Voges-Proskauer-reaktion (dvs. bildar acetylmetylkarbinol), har inget ureas, bildar inte H2S, har lysin- och ornitindekarboxylaser, men har inte arginindihydrolas. Ett karakteristiskt drag för släktet Vibrio är de flesta bakteriestammars känslighet för läkemedlet 0/129 (2,4-diamino-6,7-diazopropylpteridin), medan representanter för familjerna Pseudomonadaceae och Enterobacteriaceae är resistenta mot detta läkemedel. Vibrio är aerober och fakultativt anaerober, den optimala temperaturen för tillväxt är 18-37 C, pH 8,6-9,0 (växer i pH-intervallet 6,0-9,6), vissa arter (halofiler) växer inte i frånvaro av NaCl. G+C-halten i DNA är 40-50 mol% (för V. cholerae cirka 47 mol%). Biokemiska tester används för att differentiera inom familjen Vibrionaceae från de morfologiskt liknande släktena Aeromonas och Plesiomonas, samt för att skilja från familjen Enterobacteriaceae.
Koleravibrio-familjen skiljer sig från Pseudomonadaceae genom att den endast fermenterar glukos via Embden-Meyerhof-vägen (utan deltagande av O2), medan de förra endast konsumerar glukos i närvaro av O2. Denna skillnad mellan dem syns lätt på Hugh-Leifson-mediet. Mediet innehåller näringsagar, glukos och en indikator. Såningen görs i två kolonner med Hugh-Leifson-mediet, varav en är fylld med vaselin (för att skapa anaeroba förhållanden). Vid tillväxt av koleravibrio ändras mediets färg i båda provrören, vid tillväxt av pseudomonader - endast i provröret utan vaselin (aeroba tillväxtförhållanden).
Koleravibrion är mycket krävande för näringsmedier. Den förökar sig bra och snabbt i 1 % alkaliskt (pH 8,6-9,0) peptonvatten (PV) innehållande 0,5-1,0 % NaCl, vilket överträffar tillväxten av andra bakterier. För att hämma tillväxten av Proteus rekommenderas det att tillsätta kaliumtellurit (i en slutlig utspädning av 1:100 000) till 1 % PV. 1 % PV är det bästa anrikningsmediet för koleravibrion. Under tillväxten bildar den en delikat, lös, gråaktig film på ytan av PV:n efter 6-8 timmar, som lätt förstörs vid skakning och faller till botten i form av flingor, varvid PV:n blir måttligt grumlig. Olika selektiva medier har föreslagits för isolering av koleravibrion: alkalisk agar, gallsaltagar, alkaliskt albuminat, alkalisk agar med blod, laktos-sackaros och andra medier. Det bästa är TCBS-mediet (tiosulfatcitrat-bromotymolsackarosagar) och dess modifieringar. Oftast används dock alkalisk MPA, på vilken koleravibrion bildar släta, glasartat-transparenta, blåaktigt tonade, skivformade kolonier med viskös konsistens.
När vibrion sås genom injektion i en gelatinkolonn orsakar den efter 2 dagar vid en temperatur på 22-23 °C flytandegöring från ytan i form av en bubbla, sedan en trattformad och slutligen lager för lager.
I mjölk förökar sig vibrion snabbt, vilket orsakar koagulering efter 24-48 timmar, och sedan sker peptonisering av mjölken, och efter 3-4 dagar dör vibrion på grund av en förskjutning av mjölkens pH-värde till den sura sidan.
B. Heiberg, baserat på deras förmåga att fermentera mannos, sackaros och arabinos, delade in alla vibrioner (kolera och koleraliknande) i ett antal grupper, vars antal nu uppgår till 8.
Vibrio cholerae tillhör den första gruppen av Heiberg.
Vibrioner med liknande morfologiska, kulturella och biokemiska egenskaper som koleravibrion kallades och kallas olika: parakolera, koleraliknande, NAG-vibrioner (icke-agglutinerande vibrioner); vibrioner som inte tillhör O1-gruppen. Efternamnet betonar bäst deras släktskap med koleravibrion. Som fastställts av A. Gardner och K. Venkat-Raman har kolera- och koleraliknande vibrioner ett gemensamt H-antigen, men skiljer sig åt i O-antigener. Enligt O-antigenet är kolera- och koleraliknande vibrioner för närvarande indelade i 139 O-serogrupper, men deras antal ökar ständigt. Koleravibrion tillhör O1-gruppen. Den har ett gemensamt A-antigen och två typspecifika antigener - B och C, genom vilka tre serotyper av V. cholerae särskiljs - Ogawa-serotypen (AB), Inaba-serotypen (AC) och Hikoshima-serotypen (ABC). Koleravibrion i dissociationsstadiet har ett OR-antigen. I detta avseende används O-serum, OR-serum och typspecifika sera Inaba och Ogawa för att identifiera V. cholerae.
Åren 1992-1993 började en stor koleraepidemi i Bangladesh, Indien, Kina, Malaysia och andra länder, vars orsakande agens var en ny, tidigare okänd serovar av arten Vibrio cholerae. Den skiljer sig från V. cholerae O1 genom antigena egenskaper: den har 0139-antigenet och en polysackaridkapsel och agglutineras inte av något annat O-sera. Alla dess andra morfologiska och biologiska egenskaper, inklusive förmågan att orsaka kolera, dvs. att syntetisera exotoxin-kolerogen, visade sig likna egenskaperna hos V. cholerae O1. Följaktligen uppstod ett nytt orsakande agens för kolera, V. cholerae 0139, tydligen som ett resultat av en mutation som förändrade O-antigenet. Den fick namnet V. cholerae 0139 bengal.
Frågan om förhållandet mellan de så kallade koleraliknande vibrionerna och V. cholerae har länge varit oklar. En jämförelse av V. cholerae och koleraliknande (NAG-vibrion) med mer än 70 egenskaper avslöjade dock deras likhet på 90 %, och graden av DNA-homologi mellan V. cholerae och de studerade NAG-vibrionerna är 70–100 %. Därför kombineras koleraliknande vibrion till en art med koleravibrion, från vilken de huvudsakligen skiljer sig i sina O-antigener, i samband med vilka de kallas vibrion av icke-01-gruppen - V. cholerae non-01.
V. cholerae-arten är indelad i fyra biotyper: V. cholerae, V. eltor, V. proteus och V. albensis. El Tor-vibrion har debatterats i många år. Denna vibrio isolerades 1906 av F. Gottschlich vid karantänstationen El Tor från kroppen av en pilgrim som dog av dysenteri. F. Gottschlich isolerade flera sådana stammar. De skilde sig inte från koleravibrion i alla sina egenskaper och agglutinerades av kolera-O-serum. Eftersom det inte fanns någon kolera bland pilgrimerna vid den tiden, och långvarig bärarskap av koleravibrion ansågs osannolik, förblev frågan om V. eltors möjliga etiologiska roll i kolera kontroversiell under lång tid. Dessutom hade El Tor-vibrion, till skillnad från V. cholerae, en hemolytisk effekt. År 1937 orsakade dock denna vibrio en stor och svår koleraepidemi på ön Sulawesi (Indonesien) med en dödlighet på över 60 %. Slutligen, år 1961, blev den boven i dramat till den sjunde pandemin, och år 1962 löstes frågan om dess koleranatur äntligen. Skillnaderna mellan V. cholerae och V. eltor gäller endast vissa egenskaper. I alla andra egenskaper skiljer sig V. eltor inte fundamentalt från V. cholerae. Dessutom har det nu fastställts att biotypen V. proteus (V.finklerpriori) omfattar hela gruppen av vibriovirus, förutom gruppen 01 (och nu 0139), tidigare kallad NAG-vibriovirus. Biotypen V. albensis isolerades från Elbefloden och har förmågan att fosforescera, men efter att ha förlorat den skiljer den sig inte från V. proteus. Baserat på dessa data delas Vibrio cholerae-arten för närvarande in i fyra biotyper: V. cholerae 01 cholerae, V. cholerae eltor, V. cholerae 0139 bengal och V. cholerae non 01. De tre första tillhör två serovarer 01 och 0139. Den sista biovaren inkluderar de tidigare biotyperna V. proteus och V. albensis och representeras av många andra serovarer av vibrioner som inte agglutineras av 01- och 0139-sera, dvs. NAG-vibrioner.
Patogenicitetsfaktorer för kolera vibrio
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
Kemotaxi av Vibrio cholerae
Med hjälp av dessa egenskaper interagerar vibrion med epitelceller. Hos kolera-vibriomutanter (som har förlorat förmågan till kemotaxi) är virulensen avsevärt minskad, hos Mob-mutanter (som har förlorat rörlighet) försvinner den antingen helt eller minskar kraftigt.
Adhesions- och koloniseringsfaktorer genom vilka vibrion fäster vid mikrovilli och koloniserar tunntarmens slemhinna. Adhesionsfaktorer inkluderar mucinas, lösligt hemagglutinin/proteas, neuraminidas, etc. De främjar adhesion och kolonisering genom att förstöra ämnen som ingår i slemet. Lösligt hemagglutinin/proteas främjar separationen av vibrion från epitelcellreceptorer och deras utträde från tarmen till den yttre miljön, vilket säkerställer deras epidemiska spridning. Neuraminidas stärker bindningen mellan koleragen och epitelceller och underlättar penetrationen av toxinet in i cellerna, vilket ökar svårighetsgraden av diarré.
Koleratoxin är ett koleragen.
Så kallade nya toxiner som kan orsaka diarré, men som inte har något genetiskt eller immunologiskt samband med koleragen.
Dermoneurotiska och hemorragiska faktorer. Naturen hos dessa toxiska faktorer och deras roll i patogenesen av kolera har inte studerats tillräckligt.
[ 13 ], [ 14 ], [ 15 ], [ 16 ], [ 17 ], [ 18 ]
Endotoxiner från Vibrio cholerae
Lipopolysackarider av V. cholerae har en stark endotoxisk egenskap och orsakar allmän förgiftning av kroppen.
Den viktigaste av de listade patogenicitetsfaktorerna för kolera vibrio är exotoxinet koleragen (CTX AB), som bestämmer patogenesen för denna sjukdom. Koleramolekylen består av två fragment - A och B. Fragment A består av två peptider - A1 och A2, det har en specifik egenskap hos koleratoxin och ger det egenskaperna hos ett superantigen. Fragment B består av 5 identiska subenheter. Det utför två funktioner: 1) känner igen receptorn (monosialogangliosid) i enterocyten och binder till den; 2) bildar en intramembran hydrofob kanal för passage av subenhet A. Peptid A2 tjänar till att binda fragment A och B. Den faktiska toxiska funktionen utförs av peptid Aj (ADP-ribosyltransferas). Den interagerar med NAD, vilket orsakar dess hydrolys; Den resulterande ADP-ribosen binder till den regulatoriska subenheten av adenylatcyklas. Detta leder till hämning av GTP-hydrolys. Det resulterande GTP + adenylatcyklaskomplexet orsakar ATP-hydrolys med bildandet av cAMP. (En annan väg för cAMP-ackumulering är koleragens hämning av enzymet som hydrolyserar cAMP till 5-AMP). Manifestationen av funktionen hos ctxAB-genen som kodar för syntesen av exotoxin beror på funktionen hos ett antal andra patogenicitetsgener, särskilt tcp-generna (som kodar för syntesen av toxinkontrollerad adhesionspili - TCAP), de regulatoriska generna toxR, toxS och toxT, samt hap-generna (lösligt hemagglutinin/proteas) och neuraminidas (neuraminidas). Därför är den genetiska kontrollen av patogeniciteten hos V. cholerae komplex.
Det visade sig att det finns två patogenicitetsöar i V. cholerae-kromosomen. En av dem är genomet för den filamentösa måttligt konverterande fagen CTXφ, och den andra är genomet för den också filamentösa måttligt konverterande fagen VPIcp. Var och en av dessa patogenicitetsöar innehåller kassetter av gener specificerade i profasen, vilka bestämmer patogeniciteten hos kolerapatogenen. CTXφ-profagen bär CTX-generna, generna för de nya toxinerna zot och ace, ser-genen (adhesinsyntes) och ortU-genen (syntes av en produkt med okänd funktion). Denna kassett inkluderar också nei-genen och RS2-fagregionen, som kodar för replikation och integration av profagen i kromosomer. zot-, ace- och ortU-generna är nödvändiga för bildandet av fagvirioner när profagen exkluderas från patogenens kromosom.
VPIcp-profagen bär tcp-generna (som kodar för produktionen av pili (TCPA-protein)), toxT-, toxR- och act-gener (ytterligare koloniseringsfaktor- och mobilitetsgener (integraser och transposaser)). Transkriptionen av virulensgener regleras av tre regulatoriska gener: toxR, toxS och toxT. Dessa gener förändrar koordinerat, på transkriptionsnivå, aktiviteten hos mer än 20 virulensgener, inklusive ctxAB-, tcp- och andra gener. Den huvudsakliga regulatoriska genen är toxR-genen. Dess skada eller frånvaro leder till avirulens eller till en mer än 100-faldig minskning av produktionen av koleratoxinet CTX och TCPA. Möjligen är det så här det koordinerade uttrycket av virulensgener regleras i patogenicitetsöar som bildas av tempererat konverterande fager och i andra bakteriearter. Det har fastställts att en annan profag K139 finns i kromosomen hos V. cholerae eltor, men dess genom har studerats lite.
Hap-genen är lokaliserad på kromosomen. Således bestäms virulensen (patogeniciteten) och epidemiska kapaciteten hos V. cholerae av fyra gener: ctxAB, tcp, toxR och hap.
Olika metoder kan användas för att detektera V. choleraes förmåga att producera koleragen.
Biologiskt test på kaniner. När koleravibrioner injiceras intramuskulärt i diande kaniner (högst 2 veckor gamla) utvecklar de ett typiskt kolerasyndrom: diarré, uttorkning och kanindöd.
Direkt detektion av koleragen med PCR, IFM eller passiv immun hemolysreaktion (kolerogen binder till erytrocyternas Gmj, och de lyseras vid tillsats av antitoxiska antikroppar och komplement). Detektion av enbart förmågan att producera toxin är dock inte tillräckligt för att fastställa den epidemiska faran hos sådana stammar. För detta är det nödvändigt att detektera närvaron av hap-genen, därför är det bästa och mest tillförlitliga sättet att differentiera toxigena och epidemiska stammar av kolera vibrios av serogrupperna 01 och 0139 genom PCR med specifika primers för att detektera alla 4 patogenicitetsgener: ctxAB, tcp, toxR och hap.
Förmågan hos andra V. cholerae än serogrupperna 01 eller 0139 att orsaka sporadiska eller klusterformade diarrésjukdomar hos människor kan antingen bero på närvaron av enterotoxiner av LT- eller ST-typ, vilka stimulerar adenylat- respektive guanylatcyklassystemen, eller på närvaron av endast ctxAB-generna men ingen hap-gen.
Under den sjunde pandemin isolerades V. cholerae-stammar med varierande grad av virulens: kolerogena (virulenta), svagt kolerogena (låg virulens) och icke-kolerogena (icke-virulenta). Icke-kolerogena V. cholerae uppvisar som regel hemolytisk aktivitet, lyseras inte av den koleradiagnostiska fagen HDF(5) och orsakar inte sjukdomar hos människor.
För fagtypning av V. cholerae 01 (inklusive El Tor) föreslog S. Mukherjee uppsättningar av fager, vilka sedan kompletterades med andra fager i Ryssland. En uppsättning av sådana fager (1-7) gör det möjligt att skilja fagtyper bland V. cholerae 0116. För identifiering av toxigena och icke-toxigena V. cholerae El Tor föreslås nu i Ryssland, istället för HDF-3, HDF-4 och HDF-5, fagerna CTX* (lyserar toxigena El Tor-vibrioner) och CTX" (lyserar icke-toxigena El Tor-vibrioner).
Resistens mot kolerapatogener
Koleravibrioner överlever bra vid låga temperaturer; de förblir livskraftiga i is i upp till 1 månad; i havsvatten - upp till 47 dagar, i flodvatten - från 3-5 dagar till flera veckor, i kokt mineralvatten överlever de i mer än 1 år, i jord - från 8 dagar till 3 månader, i färsk avföring - upp till 3 dagar, på kokta produkter (ris, nudlar, kött, gröt etc.) överlever de i 2-5 dagar, på råa grönsaker - 2-4 dagar, på frukt - 1-2 dagar, i mjölk och mejeriprodukter - 5 dagar; vid förvaring i kyla ökar överlevnadsperioden med 1-3 dagar; på linnetyg förorenat med avföring överlever de i upp till 2 dagar, och på fuktigt material - en vecka. Koleravibrioner dör inom 5 minuter vid en temperatur på 80 °C, och omedelbart vid 100 °C; de är mycket känsliga för syror; de dör inom 5-15 minuter under påverkan av kloramin och andra desinfektionsmedel. De är känsliga för uttorkning och direkt solljus, men de överlever bra och länge och förökar sig till och med i öppna vattendrag och avloppsvatten rikt på organiskt material, med ett alkaliskt pH och en temperatur över 10-12 °C. De är mycket känsliga för klor: en dos aktivt klor på 0,3-0,4 mg/l vatten under 30 minuter orsakar tillförlitlig desinfektion från koleravibrioner.
Vibrios som är patogena för människor och som inte tillhör arten Vibrio Cholerae
Släktet Vibrio omfattar mer än 25 arter, varav, förutom V. cholerae, minst åtta kan orsaka sjukdom hos människor: V. parahaemolyticus, V. alginolyticus, V. vulnificus, V. fluvialis, V. fumissii, V. mimicus, V. damsela och V. hollisae. Alla dessa vibriovirus lever i hav och vikar. Infektion sker antingen genom simning eller genom att äta fisk och skaldjur. Det har visat sig att kolera- och icke-koleravibriovirus kan orsaka inte bara gastroenterit, utan även sårinfektioner. Denna förmåga har observerats i V. cholerae 01- och icke-01-grupperna, V. parahaemolyticus, V. alginolyticus, V. mimicus, V. damsela och V. vulnificus. De orsakar inflammatoriska processer i mjukvävnader när de skadas av skalet hos marina djur eller när de är i direkt kontakt med infekterat havsvatten.
Av de listade patogena icke-kolera vibrierna är de mest praktiska intressena V. parahaemolyticus, V. alginolyticus, V. vulnificus och V. fluvialis.
V. parahaemolyticus - en parahemolytisk vibrio - isolerades först i Japan 1950 under ett stort utbrott av matförgiftning orsakad av konsumtion av halvtorkade sardiner (dödligheten var 7,5%). Sjukdomsmedlet tillhörde släktet Vibrio av R. Sakazaki 1963. Han delade in de studerade stammarna i två arter: V. parahaemolyticus och V. alginolyticus. Båda arterna finns i kustnära havsvatten och hos dess invånare är de halofiler (grekiska hals - salt); till skillnad från vanliga vibrior växer halofila inte på medier utan NaCl och reproducerar sig väl vid höga koncentrationer av det. Arttillhörigheten hos halofila vibrior bestäms av deras förmåga att fermentera sackaros, bilda acetylmetylkarbinol och reproducera sig i PV med 10% NaCl. Alla dessa egenskaper är inneboende hos arten V. alginolyticus, men saknas hos V. parahaemolyticus.
Den parahemolytiska vibrion har tre typer av antigener: värmelabila flagellära H-antigener, värmestabila O-antigener som inte förstörs vid uppvärmning till 120 °C i 2 timmar, och yt-K-antigener som förstörs vid uppvärmning. Nyligen isolerade kulturer av V. parahaemolyticus har väldefinierade K-antigener som skyddar levande vibrioner från agglutination av homologa O-sera. H-antigenerna är desamma för alla stammar, men H-antigenerna hos monotrichus skiljer sig från H-antigenerna hos peritrichs. Enligt O-antigenet är V. parahaemolyticus indelad i 14 serogrupper. Inom serogrupperna är vibrioner indelade i serotyper enligt K-antigener, vars totala antal är 61. Det antigeniska schemat för V. parahaemolyticus har utvecklats endast för dess stammar isolerade från människor.
V. parahaemolyticus patogenicitet är förknippad med dess förmåga att syntetisera hemolysin, vilket har enterotoxiska egenskaper. Det senare detekteras med Kanagawa-metoden. Dess kärna ligger i det faktum att V. parahaemolyticus, patogen för människor, orsakar tydlig hemolys på blodagar innehållande 7 % NaCl. På blodagar innehållande mindre än 5 % NaCl orsakas hemolys av många stammar av V. parahaemolyticus, och på blodagar med 7 % NaCl - endast stammar med enteropatogena egenskaper. Den parahemolytiska vibrion finns vid kusterna i Japan, Kaspiska havet, Svarta havet och andra hav. Den orsakar livsmedelsburna toxiska infektioner och dysenteriliknande sjukdomar. Infektion uppstår vid intag av rå eller halvrå fisk och skaldjur infekterade med V. parahaemolyticus (havsfisk, ostron, kräftdjur etc.).
Bland de åtta icke-kolera vibriovirus som nämns ovan är den mest patogena för människor V. vulnificus, som först beskrevs 1976 som Beneckea vulnificus och sedan omklassificerades till Vibrio vulnificus 1980. Den finns ofta i havsvatten och dess invånare och orsakar olika mänskliga sjukdomar. Stammar av V. vulnificus av marint och kliniskt ursprung skiljer sig inte från varandra vare sig fenotypiskt eller genetiskt.
Sårinfektioner orsakade av V. vulnificus fortskrider snabbt och leder till bildandet av tumörer med efterföljande vävnadsnekros, åtföljda av feber, frossa, ibland svår smärta och i vissa fall amputationskrävande.
V. vulnificus har visat sig producera exotoxin. Djurförsök har visat att patogenen orsakar allvarliga lokala skador med utveckling av ödem och vävnadsnekros, följt av död. Exotoxinens roll i sjukdomens patogenes studeras.
Förutom sårinfektioner kan V. vulnificus orsaka lunginflammation hos drunkningsoffer och endometrit hos kvinnor efter exponering för havsvatten. Den allvarligaste formen av infektion orsakad av V. vulnificus är primär sepsis i samband med att man äter råa ostron (och eventuellt andra marina djur). Denna sjukdom utvecklas mycket snabbt: patienten utvecklar sjukdomskänsla, feber, frossa och utmattning, därefter svår hypotoni, vilket är den främsta dödsorsaken (dödligheten är cirka 50 %).
V. fluvialis beskrevs först som en gastroenteritpatogen år 1981. Den tillhör en undergrupp av icke-kolerapatogena vibrios som har arginindihydrolas men inga netornitin- och lysindekarboxylaser (V. fluvialis, V. furnissii, V. damsela, dvs. fenotypiskt lik Aeromonas). V. fluvialis är ett vanligt orsakande agens för gastroenterit, som åtföljs av svåra kräkningar, diarré, buksmärtor, feber och svår eller måttlig uttorkning. Den huvudsakliga patogena faktorn är enterotoxin.
Epidemiologi av kolera
Den huvudsakliga smittkällan är endast en person - en patient med kolera eller en vibriobärare, samt vatten som är förorenat med dem. Inga djur i naturen får kolera. Smittvägen är feko-oral. Smittvägar: a) den huvudsakliga - genom vatten som används för att dricka, bada och för hushållsbehov; b) kontakt-hushåll och c) genom mat. Alla större epidemier och pandemier av kolera har varit förknippade med vatten. Kolera vibrio har sådana anpassningsmekanismer som säkerställer deras populationers existens både i människokroppen och i vissa ekosystem i öppna vattendrag. Allvarlig diarré, som orsakas av kolera vibrio, leder till att tarmarna renas från konkurrerande bakterier och bidrar till den utbredda spridningen av patogenen i miljön, främst i avloppsvatten och i öppna vattendrag där de dumpas. En person med kolera utsöndrar patogenen i enorma mängder - från 100 miljoner till 1 miljard per 1 ml avföring, en vibriobärare utsöndrar 100-100 000 vibrio i 1 ml, den smittsamma dosen är cirka 1 miljon vibrio. Utsöndringstiden för koleravibrion hos friska bärare är från 7 till 42 dagar och 7-10 dagar hos dem som har tillfrisknat. Längre utsöndring är extremt sällsynt.
En egenhet hos kolera är att det efter den som regel inte sker någon långvarig smittspridning och inga stabila endemiska foci bildas. Men som redan nämnts ovan, på grund av förorening av öppna vattendrag med avloppsvatten som innehåller stora mängder organiska ämnen, tvättmedel och bordssalt, överlever koleravibrion inte bara i dem under lång tid på sommaren, utan förökar sig till och med.
Av stor epidemiologisk betydelse är det faktum att koleravibrioner av 01-gruppen, både icke-toxigena och toxigena, kan överleva under lång tid i olika akvatiska ekosystem som oodlade former. Med hjälp av polymeraskedjereaktion detekterades vct-gener av oodlade former av V. chokrae i olika vattendrag i ett antal endemiska territorier i OSS under negativa bakteriologiska studier.
Det endemiska fokuset för El Tor-koleravibrion är Indonesien, uppkomsten av denna boven i den sjunde pandemin därifrån tros vara förknippad med utvidgningen av Indonesiens ekonomiska band med omvärlden efter att landet blev självständigt, och pandemins varaktighet och blixtsnabba utveckling, särskilt dess andra våg, påverkades avgörande av bristen på immunitet mot kolera och olika sociala omvälvningar i länderna i Asien, Afrika och Amerika.
Vid kolera vidtas en rad antiepidemiska åtgärder, bland vilka de ledande och avgörande är aktiv, snabb upptäckt och isolering (sjukhusinläggning, behandling) av patienter i akuta och atypiska former och friska vibriobärare; åtgärder vidtas för att förhindra möjliga smittvägar; särskild uppmärksamhet ägnas åt vattenförsörjning (klorering av dricksvatten), efterlevnad av sanitära och hygieniska förhållanden vid livsmedelsföretag, på barninstitutioner, offentliga platser; strikt kontroll, inklusive bakteriologisk, utförs över öppna vattendrag, immunisering av befolkningen utförs etc.
Symtom på kolera
Inkubationstiden för kolera varierar från flera timmar till 6 dagar, oftast 2-3 dagar. Efter att ha kommit in i tunntarmens lumen riktas koleravibrioner, på grund av sin rörlighet och kemotaxi till slemhinnan, mot slemmet. För att tränga igenom det producerar vibrioner ett antal enzymer: neuraminidas, mucinas, proteaser, lecitinas, som förstör ämnen som finns i slemmet och underlättar vibrionernas förflyttning till epitelcellerna. Genom vidhäftning fäster vibrioner sig vid epitelets glykokalyx och, efter att ha förlorat rörlighet, börjar de föröka sig intensivt, koloniserar tunntarmens mikrovilli (se färginsats, Fig. 101.2), och producerar samtidigt en stor mängd exotoxin-kolerogen. Koleragenmolekyler binder till monosialogangliosid Gni! Och penetrerar cellmembranet, där de aktiverar adenylatcyklassystemet, och det ackumulerade cAMP orsakar hypersekretion av vätska, katjoner och anjoner Na, HCO3, Kl, Cl från enterocyter, vilket leder till koleradiarré, uttorkning och avsaltning av kroppen. Det finns tre typer av sjukdomen:
- en våldsam, svår uttorkande diarrésjukdom som leder till patientens död inom några timmar;
- mindre allvarligt förlopp, eller diarré utan uttorkning;
- asymptomatisk sjukdomsförlopp (bäring av vibrioner).
I svåra fall av kolera utvecklar patienter diarré, avföringsfrekvensen ökar, avföringen blir rikligare, blir vattnig, förlorar sin avföringslukt och ser ut som risbuljong (en grumlig vätska med slemrester och epitelceller som flyter i den). Sedan uppstår försvagande kräkningar, först av tarminnehållet, och sedan får kräkningarna utseendet av risbuljong. Patientens temperatur sjunker under det normala, huden blir blåaktig, rynkig och kall - koleraalgid. Som ett resultat av uttorkning tjocknar blodet, cyanos utvecklas, syrebrist, njurfunktionen försämras kraftigt, kramper uppstår, patienten förlorar medvetandet och döden inträffar. Dödligheten i kolera under den sjunde pandemin varierade från 1,5 % i utvecklade länder till 50 % i utvecklingsländer.
Postinfektiös immunitet är stark, långvarig och återkommande sjukdomar är sällsynta. Immuniteten är antitoxisk och antimikrobiell och orsakas av antikroppar (antitoxiner kvarstår längre än antimikrobiella antikroppar), immunminnesceller och fagocyter.
Laboratoriediagnostik av kolera
Den huvudsakliga och avgörande metoden för att diagnostisera kolera är bakteriologisk. Materialet för undersökning från patienten är avföring och kräkningar; avföring undersöks för bäring av vibriobakterier; en ligerad del av tunntarmen och gallblåsan tas för undersökning från personer som dött av kolera; från föremål i den yttre miljön undersöks oftast vatten från öppna reservoarer och avloppsvatten.
Vid utförande av en bakteriologisk undersökning måste följande tre villkor vara uppfyllda:
- så material från patienten så snabbt som möjligt (kolera vibrio överlever i avföring under en kort tid);
- Behållaren i vilken materialet tas bör inte desinficeras med kemikalier och bör inte innehålla spår av dem, eftersom koleravibrio är mycket känslig för dem;
- eliminera risken för kontaminering och infektion av andra.
Kulturen isoleras enligt följande schema: sådd på PV, samtidigt på alkalisk MPA eller något selektivt medium (TCBS är bäst). Efter 6 timmar undersöks filmen som bildats på PV, och vid behov överförs till en andra PV (såningshastigheten för koleravibrion ökar i detta fall med 10%). Från PV överförs till alkalisk MPA. Misstänkta kolonier (glasartat transparenta) överförs för att erhålla en renkultur, som identifieras genom morfologiska, kulturella, biokemiska egenskaper, motilitet och slutligen typas med hjälp av diagnostiska agglutinerande sera O-, OR-, Inaba- och Ogawa-fager (HDF). Olika alternativ för accelererad diagnostik har föreslagits, varav det bästa är den luminescerande serologiska metoden. Den möjliggör detektion av koleravibrion direkt i testmaterialet (eller efter preliminär odling i två provrör med 1 % PV, till ett av vilka kolerafagen tillsätts) inom 1,5–2 timmar. För snabb detektion av kolera vibrio har Nizhny Novgorod IEM föreslagit en uppsättning pappersindikatorskivor bestående av 13 biokemiska tester (oxidas, indol, ureas, laktos, glukos, sackaros, mannos, arabinos, mannitol, inositol, arginin, ornitin, lysin), vilka gör det möjligt att skilja representanter för släktet Vibrio från släktena Aeromonas, Plesiomonas, Pseudomonas, Comamonas och från familjen Enterobacteriaceae. För snabb detektion av kolera vibrio i avföring och i miljöobjekt kan RPGA med en antikropp diagnosticum användas. För att detektera icke-odlade former av kolera vibrio i miljöobjekt används endast polymeraskedjereaktionsmetoden.
I fall där icke-Ol-grupp V. cholerae isoleras, bör de typas med hjälp av motsvarande agglutinerande sera från andra serogrupper. Isolering av icke-Ol-grupp V. cholerae från en patient med diarré (inklusive koleraliknande diarré) kräver samma antiepidemiska åtgärder som vid isolering av Ol-grupp V. cholerae. Vid behov bestäms förekomsten av patogenicitetsgenerna ctxAB, tcp, toxR och hap i sådana vibrioner med hjälp av PCR.
Serologisk diagnostik av kolera är av hjälpkaraktär. För detta ändamål kan agglutinationsreaktionen användas, men det är bättre att bestämma titern av vibriocida antikroppar eller antitoxiner (antikroppar mot kolera bestäms med enzymimmunanalys eller immunofluorescensmetoder).
Laboratoriediagnostik av icke-kolerapatogena vibrioner
Den huvudsakliga metoden för att diagnostisera sjukdomar orsakade av icke-kolerapatogena vibriobakterier är bakteriologisk med användning av selektiva medier såsom TCBS, MacConkey, etc. Tillhörigheten av den isolerade kulturen till släktet Vibrio bestäms utifrån viktiga egenskaper hos bakterier i detta släkte.
Behandling av kolera
Behandling av kolerapatienter bör i första hand bestå av rehydrering och återställande av normal vatten-saltmetabolism. För detta ändamål rekommenderas att använda saltlösningar, till exempel med följande sammansättning: NaCl - 3,5; NaHC03 - 2,5; KCl - 1,5 och glukos - 20,0 g per 1 liter vatten. Sådan patogenetiskt underbyggd behandling i kombination med rationell antibiotikabehandling gör det möjligt att minska dödligheten vid kolera till 1 % eller mindre.
Specifikt förebyggande av kolera
För att skapa artificiell immunitet föreslogs ett koleravaccin, inklusive ett tillverkat av dödade Inaba- och Ogawa-stammar; en koleratoxoid för subkutant bruk och ett enteralt kemiskt bivalent vaccin bestående av anatoxin och somatiska antigener av Inaba- och Ogawa-serotyperna, eftersom korsimmunitet inte bildas. Emellertid är immuniteten efter vaccination inte mer än 6–8 månader, så vaccinationer utförs endast enligt epidemiologiska indikationer. Antibiotisk profylax har visat sig väl vid kolerahärdar, särskilt tetracyklin, för vilket koleravibrion är mycket känslig. Andra antibiotika som är effektiva mot V. cholerae kan användas för samma ändamål.